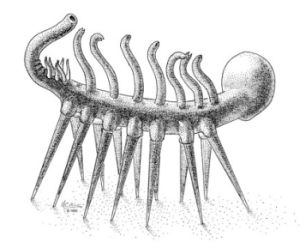
Nel vasto panorama dei Lobopodia cambriani si trovano spesso animali quantomeno bizzarri, notevolmente diversi per aspetto da qualsiasi animale vivente e che, spesso, hanno tenuto in scacco gli specialisti grazie alla loro stranezza. Basti pensare per esempio a Hallucigenia (qui il mio articolo), la cui anatomia è talmente curiosa da essere stata per quindici anni interpretata in maniera assolutamente errata. Nel vasto panorama di queste strane creature, molto probabilmenti affini ad Onychophora all'interno del superphylum Panarthropoda, si trovano pertanto organismi dalle affinità incerte; uno di questi, Diania cactiformis Liu et. al., 20011.
Un cactus semovente

Le immagini a, b, c e d rappresentano l'esemplare ELI-WD006A,B, ololotipo di Diania cactiformis, impronta, controimpronta e disegno in camera lucida; le immagini e e f raffigurano invece l'esemplare ELI-WT002A,B. La scala è di 10 millimetri. Le sigle stanno per: aba, anellamento alla base delle appendici; an, anelli; bds, base delle spine dorsali; dls, strutture a disco; fls, strutture a piega; jls,strutture simili a giunture; la, appendice sinistra; mt, tessuto muscolare; pr, proboscide; ra, appendice destra; pp, protusione posteriore;
tis, aculeo minuscolo; tu, tubercolo; wr, ruga (da http://www.nature.com/nature/journal/v470/n7335/fig_tab/nature09704_F1.html)
Si tratta di un curioso Lobopodia dall'aspetto effettivamente simile a quello di una sorta di pianta grassa con le zampe (da cui il nome specifico), proveniente dalla Heilingpu Formation, risalente al Piano 2 del Cambriano (ovvero tra i 525 e i 520 milioni di anni fa). Il tronco di questo organismo, lungo 6 centimetri, presenta dieci paia di robuste appendici, significativamente più robuste rispetto a quelle di altri Lobopodia, con la caratteristica di allargarsi distalmente fino ad essere più larghe del tronco con un diametro di oltre 2.4 millimetri. Nell'olotipo il paio di zampe più lungo sembra essere il settimo. Essendo la parte superiore del corpo pressoché uguale a quella inferiore, è difficile capire su quale lato gli esemplari si siano prevalentemente conservati. La struttura degli arti presenta delle vistose annulature, distinguibili in due morfologie: una, prossimale al corpo, dove gli anelli sono fino a 7 e più ravvicinati, ed un'altra, distale, dove invece sono fino a 15, più distanziati e presentano spine. Questi aculei sono in certi casi lunghi fino a 1.6 mm. con una base di 1 mm., rappresentando il più rimarchevole tratto di questa specie. Una delle peculiarità di questo organismo è che gli arti non sembrano terminare con un paio di artigli, come nel caso di altri Lobopodia come Hallucigenia, ma soltanto con altre spine morfologicamente identiche alle altre presenti sugli arti. All'interno degli arti è stato individuato un canale centrale, collegato al corpo e comunemente interpretato come una cavità piena di liquido avente la funzione di scheletro idrostatico, compatibile con il fatto che le appendici non sono sclerotizzate. Il tronco è una struttura cilindrica ed allungata, con una larghezza media di circa 2.4 mm. e con dei fini solchi ad anello presenti in numero di circa 4-5 ogni millimetro. Ogni anello presenta cinque o sei strutture simili a papille, in maniera non molto dissimile da Aysheaia peduncolata Walcott, 1911. Purtroppo, a differenza di altri membri di questo phylum, non è stato possibile riscontrare resti di un apparato digerente. In corrispondenza di ogni paio di arti si trovano delle strutture a disco, probabilmente degli ingrossamenti del corpo aventi la funzione di sorreggere le appendici, una caratteristica unica tra i Lobopodia sinora ritrovati. Una delle estremità del corpo è più lunga ed affusolata rispetto all'altra, tuttavia non sono stati ritrovati organi sensoriali e pertanto è difficile dire con certezza se si tratti di una proboscide oppure no.
E' difficile stabilire lo stile di vita questo organismo, tuttavia gli arti sono situati in posizione laterale, diversamente rispetto ad altri Lobopodia camminatori come Hallucigenia dove sono chiaramente siti ventralmente; inoltre, sono privi di artigli e non hanno spine sul lato inferiore. Pertanto, c'è la possibilità che Diania strisciasse ventre a terra sul fondale oceanico. L'assenza inoltre della preservazione del tratto digerente, a causa della mancanza di sedimenti al suo interno, porta a ritenere che si trattasse di un saprofago.
Questioni da piante grasse
L'analisi cladistica, basata su 42 caratteristiche e 27 taxa, è molto interessante e suscita scalpore perché non solo colloca Diania alla base di un gruppo di Lobopodia cambriani, ma anche perché sembra sconvolgere la monofilia di questo gruppo e le presunte strette relazioni di Diania con Arthropoda.

Infatti, la peculiarità di questa analisi è l'identificazione di tre gruppi all'interno di Panarthropoda: il primo è costituito dagli Onychophora e dai Tardigrada odierni più alcune forme fossili (incluso Aysheaia), il secondo da diversi Lobopodia cambriani incluso Diania (che risulta essere una forma basale) il terzo infine sarebbe un gruppo parafiletico includente tra gli altri Megadictyon (il quale non è imparentato in maniera più o meno stretta con Hallucigenia, tra l'altro), Dinocaridida ed Arthropoda. Questo studio contraddice anche l'analisi cladistica pubblicata su Nature nel 2011, la quale invece presentava questo organismo come una sorta di anello di congiunzione tra i Dinocaridida e gli Arthropoda. Pertanto, è difficile dire quali siano le affinità più prossime di Diania, e sorgono da questo studio nuove problematiche inerenti la monofilia di questo gruppo di misteriosi misteriosi animali.
BIBLIOGRAFIA
Liu, Steiner, Dunlop, Keupp, Shu, Ou, Han, Zhang & Zhang, "An armoured Cambrian lobopodian from China with arthropod-like appendages", Nature 470, pp. 526-530, 2011
Ma, Edgecombe, Legg & Hou, "The morphology and phylogenetic position of the Cambrian lobopodian Diania cactiformis", Journal of Systematic Palaeontology, 2013
E' difficile stabilire lo stile di vita questo organismo, tuttavia gli arti sono situati in posizione laterale, diversamente rispetto ad altri Lobopodia camminatori come Hallucigenia dove sono chiaramente siti ventralmente; inoltre, sono privi di artigli e non hanno spine sul lato inferiore. Pertanto, c'è la possibilità che Diania strisciasse ventre a terra sul fondale oceanico. L'assenza inoltre della preservazione del tratto digerente, a causa della mancanza di sedimenti al suo interno, porta a ritenere che si trattasse di un saprofago.
Questioni da piante grasse
L'analisi cladistica, basata su 42 caratteristiche e 27 taxa, è molto interessante e suscita scalpore perché non solo colloca Diania alla base di un gruppo di Lobopodia cambriani, ma anche perché sembra sconvolgere la monofilia di questo gruppo e le presunte strette relazioni di Diania con Arthropoda.
Il cladogramma: Diania sarebbe alla base di uno dei tre distinti filoni in cui si suddividerebbe Panarthropoda (da Ma et al., vedi bibliografia)
Infatti, la peculiarità di questa analisi è l'identificazione di tre gruppi all'interno di Panarthropoda: il primo è costituito dagli Onychophora e dai Tardigrada odierni più alcune forme fossili (incluso Aysheaia), il secondo da diversi Lobopodia cambriani incluso Diania (che risulta essere una forma basale) il terzo infine sarebbe un gruppo parafiletico includente tra gli altri Megadictyon (il quale non è imparentato in maniera più o meno stretta con Hallucigenia, tra l'altro), Dinocaridida ed Arthropoda. Questo studio contraddice anche l'analisi cladistica pubblicata su Nature nel 2011, la quale invece presentava questo organismo come una sorta di anello di congiunzione tra i Dinocaridida e gli Arthropoda. Pertanto, è difficile dire quali siano le affinità più prossime di Diania, e sorgono da questo studio nuove problematiche inerenti la monofilia di questo gruppo di misteriosi misteriosi animali.
BIBLIOGRAFIA
Liu, Steiner, Dunlop, Keupp, Shu, Ou, Han, Zhang & Zhang, "An armoured Cambrian lobopodian from China with arthropod-like appendages", Nature 470, pp. 526-530, 2011
Ma, Edgecombe, Legg & Hou, "The morphology and phylogenetic position of the Cambrian lobopodian Diania cactiformis", Journal of Systematic Palaeontology, 2013