Con questo post si conclude la quadrilogia sulle "stranezze cambriane": dopo aver affrontato gli strani casi riguardanti Nectocaris, gli Anomalocarididae e curiosi Mollusca (o organismi che assomigliano a dei Mollusca) del Paleozoico e le loro curiose interpretazioni storiche, derivanti da fossili incompleti e incompresi, è ora di occuparsi dell'animale che forse più di tutti rappresenta degnamente l'intera categoria: si tratta di Hallucigenia sparsa Conway Morris, 1977.
Un fossile allucinante
Quando il professor Conway Morris descrisse il minuscolo olotipo (si parla di neppure 0,3 centimetri), si accorse fin da subito che si trattava di qualcosa che non assomigliava a niente di conosciuto. L'esemplare infatti presentava due paia di lunghe spine ed una curiosa serie di sette tentacoli artigliati dalla parte opposta; inoltre, la controimpronta mostrava una strana struttura rotondeggiante, vagamente simile ad una regione cefalica ma apparentemente priva di organi di senso. Come se non bastasse, la regione (apparentemente) terminale del corpo si incurvava in una sorta di "coda", con alla base altre due paia di tentacoli rivolti verso l'alto. L'organismo che ne risultava, in effetti, era decisamente allucinante, come indicava il suo nome. L'unica spiegazione che fu proposta era che le spine in realtà fossero delle zampe lunghe e rigide, simili a trampoli, sfruttate da Hallucigenia per camminare sui fondali fangosi dove viveva.

USNM 83935, l'olotipo di Hallucigenia sparsa; impronta e controimpronta, sotto luce polarizzata o meno ( da http://burgess-shale.rom.on.ca/en/fossil-gallery/view-species.php?id=60&m=3&)
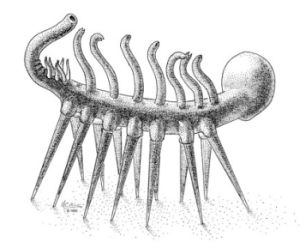
Una immagine storica di Hallucigenia, di cui è possibile ammirarne tutta la stranezza (da http://palaeos.com/metazoa/ecdysozoa/panarthropoda/hallucigeniidae.html)
Tuttavia, anche questa interpretazione lasciava adito a diversi dubbi. Anzitutto, Hallucigenia era realmente un organismo singolo o si trattava dell'appendice raptatoria di un animale ben più grande, come nel caso di Anomalocaris? Supponendo però la sua "indipendenza", in quale gruppo di animali doveva essere collocato? Nessun phylum di organismi viventi sembrava adatto allo scopo: il cephalon globulare presente in alcuni esemplari (ma apparentemente privo di organi di senso), le due file di spine, i tentacoli dorsali e lo strano "addome" tendente verso l'alto non lasciavano intravvedere somiglianze evidenti con nessun altro gruppo, moderno o estinto. Addirittura i tentacoli furono interpretati persino come un serie di proboscidi boccali, collegate ad un tubo digerente centrale e che pertanto avrebbero indicato che o Hallucigenia fosse un organismo coloniale, o persino l'animale più strano mai esistito, dotato di un sistema di bocche multiple. Il mistero ha continuato a restare tale per oltre dieci anni, finché non venne finalmente ritrovato un fossile nell' Amgan Stage, negli Stati Uniti, che permise di iniziare a capire come stessero realmente le cose.
Sopra è sotto
La svolta fu il ritrovamento e la descrizione di Microdictyon robisoni Bengtson, Matthews et Missarzhevsky, 1986. Questo piccolo organismo fu solo il primo membro di questo prolifico genere di animali cambriani, caratterizzati dalla presenza di dieci paia di scleriti e di diverse zampe tubulari, con quattro tentacoli più corti nella regione posteriore situati ventralmente. La somiglianza di questi piccoli organismi con Hallucigenia era sospetta, ed il sospetto diventò certezza quando, nel 1992, furono trovate traccia di una seconda fila di tentacoli, che si rivelarono pertanto essere in realtà zampe. Anche questa "stranezza cambriana" era stata vittima delle sue piccole dimensioni (si parla di una lunghezza massima, nelle tre specie di Hallucigenia sinora descritte, di circa 3 centimetri), della sua relativa rarità e del cattivo stato di conservazione in cui versavano i reperti. Pertanto, con questi nuovi dati era possibile ribaltare il piccolo animale, e le spine divenivano finalmente degli strumenti atti a difendersi ed i tentacoli dei veri e propri mezzi di locomozione. Addirittura, la somiglianza relativa con i moderni Onycophora portò ad ipotizzare che si trattasse di un rappresentate di questo gruppo. Infine, la struttura globulare si trovava realmente nella porzione anteriore, però come hanno mostrato anche alcuni recenti ritrovamenti del Chengjiang non era priva di organi di senso (sono stati ritrovati gli occhi), né tantomeno così arrotondata; quella forma anomala era in realtà dovuta allo schiacciamento durante la fossilizzazione ed alla decomposizione.
La svolta fu il ritrovamento e la descrizione di Microdictyon robisoni Bengtson, Matthews et Missarzhevsky, 1986. Questo piccolo organismo fu solo il primo membro di questo prolifico genere di animali cambriani, caratterizzati dalla presenza di dieci paia di scleriti e di diverse zampe tubulari, con quattro tentacoli più corti nella regione posteriore situati ventralmente. La somiglianza di questi piccoli organismi con Hallucigenia era sospetta, ed il sospetto diventò certezza quando, nel 1992, furono trovate traccia di una seconda fila di tentacoli, che si rivelarono pertanto essere in realtà zampe. Anche questa "stranezza cambriana" era stata vittima delle sue piccole dimensioni (si parla di una lunghezza massima, nelle tre specie di Hallucigenia sinora descritte, di circa 3 centimetri), della sua relativa rarità e del cattivo stato di conservazione in cui versavano i reperti. Pertanto, con questi nuovi dati era possibile ribaltare il piccolo animale, e le spine divenivano finalmente degli strumenti atti a difendersi ed i tentacoli dei veri e propri mezzi di locomozione. Addirittura, la somiglianza relativa con i moderni Onycophora portò ad ipotizzare che si trattasse di un rappresentate di questo gruppo. Infine, la struttura globulare si trovava realmente nella porzione anteriore, però come hanno mostrato anche alcuni recenti ritrovamenti del Chengjiang non era priva di organi di senso (sono stati ritrovati gli occhi), né tantomeno così arrotondata; quella forma anomala era in realtà dovuta allo schiacciamento durante la fossilizzazione ed alla decomposizione.
Realmente un onicoforo?
Tuttavia, nonostante l'organismo sia stato messo nella giusta posizione, non pochi dubbi permangono attorno ad esso. Hallucigenia era realmente un Onycophora? In molti ritengono che lei, Microdictyon e simili altri organismi del Cambriano siano da radunarsi in un phylum a parte chiamato Lobopoda. Secondo altri, invece, Onycophora sarebbe da intendersi come un taxon interno a Lobopoda o un suo sister group. Addirittura, c'è la possibilità che altri enigmatici gruppi del Cambriano come Dinocaridida possano anch'essi essere collocati all'interno di questo gruppo. Le tesi sono molteplici, ma la reale posizione sistematica di Lobopodia (ed il suo valore) restano sostanzialmente incerti. Segno, anche questo, che abbiamo appena incominciato ad interrogarci su quali siano i reali enigmi cambriani.
BIBLIOGRAFIA
Conway Morris, "A new metazoan from the Cambrian Burgess Shale of British Columbia", Paleontology, vol. 20, pp. 623-640
Bengtson, Matthews & Missarzhevsky, "The Cambrian Net fossil Microdictyon", Problematic Fossil Taxa, vol. 5, pp. 97-115, 1986
Ramsk"The second leg row of Hallucigenia discovered", Lethaia, vol. 25, pp. 221-224, 1992
Steiner, Hu, Liu & Keupp, "A new species of Hallucigenia from the Cambrian Stage Wulongqing Formation of Yunnan (South China) and the structure of sclerites in lobopodians", Bulletin of Geoscience, vol. 87, pp. 107-124, 2012
Ma et al., "Morphology of Cambrian lobopodian eyes from the Chengjiang Lagerstätte and their evolutionary significance", Arthropod Structure and Development, vol. 41, pp. 495-504, 2012
Tuttavia, nonostante l'organismo sia stato messo nella giusta posizione, non pochi dubbi permangono attorno ad esso. Hallucigenia era realmente un Onycophora? In molti ritengono che lei, Microdictyon e simili altri organismi del Cambriano siano da radunarsi in un phylum a parte chiamato Lobopoda. Secondo altri, invece, Onycophora sarebbe da intendersi come un taxon interno a Lobopoda o un suo sister group. Addirittura, c'è la possibilità che altri enigmatici gruppi del Cambriano come Dinocaridida possano anch'essi essere collocati all'interno di questo gruppo. Le tesi sono molteplici, ma la reale posizione sistematica di Lobopodia (ed il suo valore) restano sostanzialmente incerti. Segno, anche questo, che abbiamo appena incominciato ad interrogarci su quali siano i reali enigmi cambriani.
BIBLIOGRAFIA
Conway Morris, "A new metazoan from the Cambrian Burgess Shale of British Columbia", Paleontology, vol. 20, pp. 623-640
Bengtson, Matthews & Missarzhevsky, "The Cambrian Net fossil Microdictyon", Problematic Fossil Taxa, vol. 5, pp. 97-115, 1986
Ramsk"The second leg row of Hallucigenia discovered", Lethaia, vol. 25, pp. 221-224, 1992
Steiner, Hu, Liu & Keupp, "A new species of Hallucigenia from the Cambrian Stage Wulongqing Formation of Yunnan (South China) and the structure of sclerites in lobopodians", Bulletin of Geoscience, vol. 87, pp. 107-124, 2012
Ma et al., "Morphology of Cambrian lobopodian eyes from the Chengjiang Lagerstätte and their evolutionary significance", Arthropod Structure and Development, vol. 41, pp. 495-504, 2012













